
Květ

Květ je orgánem krytosemenných rostlin. Je omezeného vzrůstu a jeho části se přímo nebo nepřímo podílejí na pohlavním rozmnožování. Nepřímou úlohu při rozmnožování mají květní obaly, které mohou lákat opylovače; rozmnožování se přímo účastní pohlavní orgány: tyčinky a pestíky.
Květy krytosemenných se skládají z těchto částí (ne všechny ale musejí být přítomné): nerozlišené květní obaly neboli okvětí, nebo květní obaly rozlišené na kalich a korunu. Dále se v květu může nacházet soubor tyčinek (andreceum) a pestíků (gyneceum). Součásti květu jsou umístěné na květním lůžku. Květ ke stonku připojuje květní stopka.
Tyčinky vytvářejí pyl, který se dostává na bliznu pestíku. Vyklíčí v pylovou láčku a oplodní vajíčko v semeníku, přesněji vaječnou buňku, která se ve vajíčku nachází. Z oplodněné vaječné buňky vznikne zárodek nové rostliny, z vajíčka vznikne semeno a ze semeníku (a případně dalších částí květu) se stane plod.
Květy obsahující pouze pestíky se označují jako samičí, květy se samotnými tyčinkami se popisují jako samčí. Pokud se v květu nacházejí tyčinky i pestíky, květ je oboupohlavný.


Definice
[editovat | editovat zdroj]Květy jsou typické pro krytosemenné (kvetoucí[1]) rostliny. Květ může být definován jako „kompaktní, oboupohlavný rozmnožovací výhon omezeného vzrůstu obsahující megasporangia (plodolisty), mikrosporangia (tyčinky) a sterilní květní obaly, které obsahují aspoň jeden plochý neplodný orgán“. Tato definice nemůže zcela vystihnout všechny květy, protože druhotně může docházet ke ztrátě určitých částí.[2]
Některé definice (zvláště v německojazyčném prostředí) za květ považují i rozmnožovací orgány nahosemenných rostlin.[3][4] Ty je však přesnější označovat jako šištice. Tento článek květ popisuje pouze jako orgán krytosemenných rostlin.
Morfologie
[editovat | editovat zdroj]Jednotlivé součásti květu jsou na zkráceném výhonu uspořádány odspoda nahoru tak, že výše umístěné orgány se nacházejí více uvnitř. Toto rozložení květních částí odpovídá pořadí jejich vývoje.
Květní obaly
[editovat | editovat zdroj]Květní obaly jsou sterilní ploché útvary, které jsou nedílnou součástí květu. Pokud všechny květní obaly vypadají stejně, jedná se o květ stejnoobalný (homochlamydeický) a jeho obaly se označují jako okvětí (například u tulipánu). Okvětí se může skládat z jednoho či dvou a více kruhů okvětních lístků (tepala), případně mohou být okvětní lístky uspořádané ve spirále.
Květní obaly mohou být i rozlišené (heterochlamydeické). V takovém případě jsou vnější lístky většinou zelené a označují se jako kališní lístky (sepala), které tvoří kalich (calyx). Vnitřní lístky jsou obvykle pestrobarevné, označují se jako korunní lístky (petala) a tvoří korunu (corolla). Květní obaly mohou být nesrostlé (volné), nebo mohou srůstat, a to sice kongenitálně (při samotném vzniku) nebo postgenitálně (druhotně). Květní obaly vznikly v průběhu evoluce z listů nebo tyčinek.[5]
Tyčinky
[editovat | editovat zdroj]
Mikrosporofyly krytosemenných se označují jako tyčinky. Ty vytvářejí mikrospory neboli pyl. Tyčinka se skládá z prašníku, který je umístěný na nitce. Prašník se skládá obvykle ze dvou prašných pouzder, která jsou srostlá spojidlem (konektivem). Prašné pouzdro obsahuje obvykle dva prašné váčky. Soubor tyčinek v květu se označuje jako andreceum. V jediném květu se může nacházet až kolem 2000 tyčinek, ale může v něm také být tyčinka jediná.
Pestíky
[editovat | editovat zdroj]U krytosemenných rostlin jsou vajíčka ukryta v semeníku, který je součástí pestíku. Soubor pestíků se nazývá gyneceum. Semeník obsahuje placenty, na nichž jsou umístěna vajíčka. Na semeník může navazovat čnělka, na jejímž vrcholu je blizna. Na bliznu usedá pyl. Blizna je s vajíčky spojena zvláštním pletivem, které dovoluje prorůstání pylové láčky. Pestík se může skládat z jediného plodolistu (apokarpní gyneceum), nebo může srůstat z více plodolistů (cenokarpní gyneceum). Pestíků vzniklých z jednoho plodolistu může být v květu větší množství. Květy některých rostlin mohou být sterilní (tedy mají redukované tyčinky a pestíky) a sloužit jen k lákání opylovačů na jiné květy, jako např. některé jazykovité květy v úborech hvězdnicovitých.
Květní lůžko
[editovat | editovat zdroj]
Na květním lůžku (receptaculum, torus) se nacházejí další součásti květu. Květní lůžko vzniká rozšířením vzrostného vrcholu stonku (květní stopky), může být ploché, miskovitě prohloubené či vyklenuté.[5]
Květní lůžko může být protažené od středu květu. Příkladem je trubkovitý květ hvězdnicovitých nebo květy mnoha tykvovitých. Rozšířený útvar tvořený květním lůžkem a bázemi dalších částí květu se označuje jako češule (hypanthium). Pokud se semeník nachází v češuli a zhruba v jeho polovině z okraje češule vyrůstají květní obaly a tyčinky, označuje se jako polospodní. Pokud je celý semeník vnořen do češule a květní obaly začínají nad ním, označuje se jako semeník spodní.
Květní lůžko může oddáleně od dalších částí květu nést určité orgány: gyneceum (poté se označuje jako gynofor, např. u kapary trnité), gyneceum a andreceum (androgynofor, např. u mučenek), nebo gyneceum, andreceum a korunu (anthofor, např. u čeledi hvozdíkovité).
Nektaria
[editovat | editovat zdroj]Nektaria (medníky) jsou žlázy, které produkují nektar jako odměnu pro opylovače. Nektar obsahuje zejména cukry, ale může obsahovat také proteiny, aminokyseliny a vitamíny. Někdy je v něm i podíl olejů (např. u drchničky). Nektaria se mohou nacházet na květních obalech, tyčinkách, pestících, květním lůžku i vně samotného květu (pak se označují jako extraflorální nektária).
Listeny a listence
[editovat | editovat zdroj]Květ či květenství může vyrůstat v paždí listenu. Na květní stopce se mohou nacházet i listence, často v počtu dvou. Listeny a listence vznikají metamorfózou listu; nejsou tedy přímo součástí květu, ale souvisejí s ním. U některých rostlin jsou květenství nebo květy nenápadné, kdežto listeny jsou výrazně zbarvené a přejímají funkci lákání opylovačů (např. u pryšce nádherného, toulec u árónovitých rostlin). U některých skupin rostlin listen chybí, jako např. u brukvovitých.[6]
Poloha a uspořádání součástí
[editovat | editovat zdroj]Uspořádání součástí květu na květním lůžku se označuje jako fylotaxe. Části květu mohou být uspořádané ve spirálách (acyklické uspořádání) nebo v kruzích (cyklické uspořádání). Spirální uspořádání součástí květů se považuje za evolučně původnější a často se vyskytuje v květech, které sestávají z více částí. Úhel mezi dvěma sousedními orgány se blíží 137,5°, což odpovídá zlatému řezu.
Mnohdy jsou šroubovitě uspořádány jen vnější orgány, vnitřní orgány jsou uspořádány v kruzích (přeslenech). Takové uspořádání se označuje jako spirocyklické. Náznaky přechodu ze spirálního na cyklické uspořádání lze pozorovat např. u magnólie (Magnolia denudata), kdy po vytvoření třech okvětních lístků dochází k pozastavení růstu a lístky jsou tak uspořádány v kruzích po třech. I u cyklických květů se lístky někdy vyvíjejí ve spirále, ale jsou tak blízko sebe, že se jejich uspořádání považuje za kruhové.
V každém kruhu se nachází jenom jeden typ orgánů. Cyklické uspořádání květu je také spojeno se zmenšením počtu jeho částí. Většina cyklicky uspořádaných oboupohlavných květů obsahuje 5 kruhů orgánů: kruh kališních lístků, kruh korunních lístků, 1–2 kruhy tyčinek a 1 kruh plodolistů. U většiny krytosemenných je počet orgánů v kruhu pevně daný, poté se hovoří o dvou-, troj-, čtyř- či pětičetných květech. Četnost květu tak může sloužit jako určovací znak.
Orgány v sousedních kruzích jsou obvykle uspořádané střídavě, tedy orgán jednoho kruhu se nachází mezi orgány kruhu druhého. U pětičetných květů je někdy vnitřní kruh tyčinek vytlačen semeníkem směrem od středu květu. Tento jev se nazývá obdiplostemonie. Jedna součást květu se tak může nacházet přímo před jinou součástí náležící do sousedního kruhu.
Souměrnost květu
[editovat | editovat zdroj]Pokud lze květem proložit tři a více rovin souměrnosti, jedná se o květ pravidelný (aktinomorfní, např. u tulipánu). Květ se dvěma rovinami souměrnosti je bisymetrický (jako např. u srdcovky). Jednu rovinu souměrnosti mají souměrné (zygomorfní) květy. Tato rovina obvykle prochází středem květu (od listenu k mateřskému stonku), jako např. u hluchavkovitých, nebo poblíž středu (jako u dymnivky). Asymetrické květy (např. u některých třezalek) žádnou rovinu souměrnosti nemají, tak tomu je i u spirálně uspořádaných květů. Asymetrické květy někdy bývají uspořádané v symetrických dvojicích, což je pravděpodobně výhodnější pro lákání opylovačů.[7]
Stavba a souměrnost květu lze vyjádřit grafickými květními diagramy nebo textovými květními vzorci.
Větší množství květů bývá uspořádáno do květenství.

Pohlavnost květů
[editovat | editovat zdroj]Pro krytosemenné rostliny jsou typické oboupohlavné květy. Přesto existují i květy jednopohlavné, v kterých jsou orgány druhého pohlaví buď zakrnělé, nebo zcela chybí. Jednopohlavné květy mohou být tyčinkové (samčí) nebo pestíkové (samičí). Rostliny mohou být jednodomé, tedy se u nich vyskytují oddělené samčí a samičí květy (v případě jiných skupin rostlin např. i šištice) na jedné rostlině. U dvoudomých rostlin se jednopohlavné květy nacházejí odděleně na různých rostlinách. Některé rostliny mohou mít jak oboupohlavné, tak jednopohlavné květy na jednom jedinci.
Vyvolání kvetení
[editovat | editovat zdroj]Indukce kvetení je jev, kdy meristémy vegetativních orgánů začnou vytvářet květy. To může být ovlivněno vnějšími (endogenními) či vnějšími (exogenními) faktory. Indukce kvetení je nevratná, nelze ji ani zastavit. Ovlivňují ji čtyři důležité typy signálů:
- Na vyvolání kvetení se podílejí vnitřní neboli autonomní faktory, ale není o nich mnoho známo. Velké množství druhů rostlin, zejména stromů, před zahájením tvorby květů potřebuje dosáhnout určitého věku či dorůst určité velikosti. U hrachu je doba kvetení určena geneticky. Pozdní ekotypy huseníčku rolního (Arabidopsis thaliana) vytváří od určitého věku květy samy, aniž by došlo k indukci kvetení faktory prostředí. Aktivují se u něj heterochronní geny jako např. FCA (Flowering Control Adabidopsis).
- U většiny rostlin je doba kvetení určena vnějšími faktory. Mezi nejdůležitější z nich patří délka dne a období chladu (jarovizace neboli vernalizace). Dle potřeby denního světla se rostliny dělí na krátko- a dlouhodenní. Jako receptor světla působí systém fytochromů v listu, konkrétně protein zvaný kryptochrom. Také protein FKF1 slouží jako fotoreceptor a reaguje na délku dne. Tyto systémy ovlivňují gen CO, jehož exprese sama o sobě vede k zahájení kvetení. Přenášení signálu z listů do květů se přisuzovalo hormonu florigenu, ale jeho identita byla dlouho neznámá. Je možné, že je vytvářen díky genu FT, jehož transkripce probíhá v listech, ale přesto ovlivňuje květní meristémy.[8][9]
- Zvláště monokarpní rostliny (takové, které kvetou jen jednou za život) vyžadují vernalizaci, delší periodu chladu, aby mohly vykvést. Jsou známy geny aktivované chladem, např. Frigida (FRI) u Arabidopsis a VRN1 a VRN2 u ozimé pšenice. Látky vytvořené na jejich základě inhibují gen FLC, který potlačuje kvetení. Celkově se geny FRI nebo VRN podílejí na tvorbě květů, i když ji přímo nespouští. Ve většině případů ve vyvolání kvetení hrají roli i další faktory.
- Ke stimulaci kvetení se v nepřítomnosti dlouhých dnů dají využít rostlinné hormony, např. giberelin, ale mechanizmus jejich fungování ještě není znám.
Vznik květu
[editovat | editovat zdroj]Díky heterochronním genům se z meristémů vegetativních orgánů začnou vytvářet orgány generativní, samotný vznik květu ovlivňuje více genů, které spolu interagují.
Geny pro symetrii
[editovat | editovat zdroj]Tyto geny ovlivňují jak strukturu květu a jeho symetrii, tak počet květních orgánů a květů v květenství. Mutace těchto genů pozmění stavbu květu, tyto geny jsou často pojmenovány na základě toho, jak se jejich mutace projeví. Mutace genu Cycloidea (CYC) nebo Dichotoma (DICH) u hledíku dá vzniknout pravidelným květům místo obvyklých květů souměrných. Takové mutace se odehrávají i v přírodě; jev, kdy místo souměrného květu vznikne květ pravidelný se nazývá pelorie. Mutace genu Cauliflower (CAL) u huseníčku nebo brokolice vede k vytváření rozvětveného květenství bez funkčních květů. CAL inhibuje gen TFL1, který se podílí na rozvětvování květenství a stavbě výhonků. Jedinci se zmutovaným genem TFL1 vytvářejí květenství vždy jen s jedním květem.

Geny ovlivňující identitu květu
[editovat | editovat zdroj]Během rozvoje stonků květenství se vegetativní meristémy mění na meristémy vytvářející květy díky expresi genů Leafy (LFY) a Apetala1 (AP1). Oba nesou informaci o podobě transkripčních faktorů a náleží do skupiny genů, které integrují signály vedoucí ke kvetení. Funkce těchto a podobných genů se zkoumala na huseníčku, hledíku a křížencích petúnií. Nejdůležitější interakce jsou následující:
- FLC (Flowering Locus C) integruje signály o vernalizaci s autonomními signály. FLC je transkripční faktor, zabránění jeho činnosti vede k dřívějšímu kvetení. Množství proteinu FLC/odpovídající mRNA koreluje s dobou kvetení. FLC potlačuje aktivátor kvetení SOC1.
- SOC1 (Suppressor of Overexpression of Constans) je aktivován prostřednictvím giberelinů a CO souvisejícího s dlouhým dnem, potlačuje ho FLC. Samotné zrušení potlačení FLC nevede k aktivaci SOC1, tento gen musí být aktivován přes CO nebo gibereliny.
- LFY se podobně jako SOC1 aktivuje gibereliny nebo dlouhým dnem. K tomu dochází prostřednictvím SOC1 nebo přes Agamous-Like24 (AGL24).
- FT (Flowering Locus T) je aktivován dlouhým dnem nebo přes CO, inhibuje ho FLC. FT aktivuje AP1.
Dva poslední geny v této návaznosti jsou LFY a AP1, takzvané geny identity květního meristému. Ty spolu interagují. AP1 se aktivuje prostřednictvím LFY; sice vytvoří květ i v absenci LFY, ale ne se správnými orgány. Již zmíněný TFL1 na jedné a LFY a AP1 na straně druhé se navzájem potlačují. TFL1 se používá k zabránění předčasného kvetení.
Vytváření orgánů květu
[editovat | editovat zdroj]
Jednotlivé součásti květu se vytvářejí podle tzv. modelu ABC. Jsou tři skupiny genů, které ovlivňují interakci mezi květními orgány:
- geny A ovlivňují kalich
- geny A a B ovlivňují korunu
- B a C ovlivňují tyčinky
- C je zodpovědný za stavbu plodolistů
Geny A a C se navzájem potlačují, takže nedochází k překrývání jejich působení. Pokud jeden z nich chybí, druhý se projevuje v celém květu. Model ABC v roce 1991 navrhli E. Coen a E. Meyerowitz[10], od té doby byl mnohokrát potvrzen a rozšířen.
U huseníčku jsou známy následující geny:
- třída A: AP1 a AP2
- třída B: AP3 a Pistillata (PI)
- třída C: Agamous (AG)
Exprese těchto genů v orgánech neprobíhá jen na začátku růstu, ale celou dobu. AP1 neovlivňuje jen květní meristém, ale také identitu orgánů. V rané fázi vývoje květu se nachází v celém meristému, poté jen ve květních obalech. Mutace tohoto genu vedou ke změně identity orgánů. Pokud nefunguje gen A, místo okvětí se vyvíjí plodolisty a tyčinky (neboť probíhá exprese genu C). Pokud jsou zmutovány všechny třídy genů, vytvářejí se místo květních součástí listy. Květy jedinců se zmutovanými geny C neomezeně rostou.
Dodatečně byla objevena čtvrtá třída genů, E, která je u Arabidopsis zastoupena čtyřmi geny Sepallata1 až 4 (SEP). Jsou exprimovány při stavbě korunních lístků, tyčinek a plodolistů a pro podobu orgánů jsou taktéž nezbytné. Trojnásobní mutanti (SEP1 až 3) vytvářejí jen kalich, čtyřnásobní mutanti jen listy.[2]
Pátá třída genů, D, je zodpovědná za podobu vajíček. U petúnií se jedná o geny Floral Binding Protein (FBP) 7 a 11. Dvojnásobní mutanti vytvářejí místo vajíček struktury podobné plodolistům. U Arabidopsis podobnou úlohu mají geny Seedstick (STK), Shatterproof1 (SHP1) a SHP2.
Převládající část genů zodpovědných za podobu rostlinných orgánů patří do skupiny MADS-Box genů. U rostlin jsou MADS-Box geny značně rozšířeny. Mimo jiné jsou součástí rostlinných homeotických genů (jako AGAMOUS nebo DEFICIENS), které se podílejí na určování výstavby rostlinných orgánů včetně květu. Příkladem jejich funkce je uspořádání orgánů v květu. Pomocí vnější aktivace těchto genů se dosáhlo tvorby květů z vegetativních orgánů.
Úloha genů zodpovědných za identitu květu, jejichž exprese probíhá v celém květním meristému, se teprve začíná objasňovat. Zdá se, že k expresi genu třídy B (AP3) dochází kombinací LFY, AP1 a genu Unusual Flower Organs (UFO). LFY se aktivuje s dalším koaktivátorem, Wuchsel (WUS), a společně vedou k expresi genu AG, který náleží do třídy C. Po aktivaci gen AG zablokuje gen WUS. To zastaví růst květní stopky a vytvoří květní lůžko.
Další krok k pochopení problematiky přinesl Thießes (2001), který navrhl tzv. kvartet model. Podstata tohoto modelu spočívá v tom, pro vznik každého určitého orgánu je potřeba konkrétní komplex čtyř proteinů. Pro každý orgán existuje aspoň jeden specifický tetramer. Toto zatím bylo prokázáno u huseníčku. Zmíněný model slouží jako základ mnoha dalších prací.[2]
Variace na model ABC
[editovat | editovat zdroj]
Srovnávací studie řady druhů rostlin ukázaly, že model ABC ve výše popsané podobě platí jen u vyšších dvouděložných rostlin. U vývojově původnějších skupin krytosemenných je stavba květu rozmanitější. U stulíků (Nuphar) se geny neprojevují zcela ohraničeně, takže přechod od okvětních lístků napodobujících kalich k okvětním lístkům, tyčinkám a pestíkům je pozvolný.[2]
Tulipán má okvětní lístky ve dvou kruzích. V rámci obou kruhů se projevují geny třídy B.[2]
Lipnicovité rostliny mají oproti dalším rostlinám dost pozměněnou stavbu květu, nicméně u nich funguje model ABC. Pluška je tvořena za účasti genů A a osiny prostřednictvím genů A a B, takže jsou homologické koruně a kalichu.[2]
Šťovík (Rumex) vytváří redukované a málo nápadné květní obaly. V jeho květech jsou geny skupiny B zodpovědné za tvorbu tyčinek, a na stavbě květních obalů se podílejí pouze geny skupiny A.
-
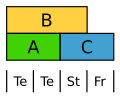 Varianta ABC modelu u tulipánu. Te = okvětní lístky.
Varianta ABC modelu u tulipánu. Te = okvětní lístky. -
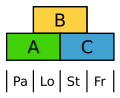 Model ABC u rýže. Pa = pluška, Lo = plenky.
Model ABC u rýže. Pa = pluška, Lo = plenky. -
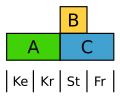 Varianta modelu ABC u šťovíku.
Varianta modelu ABC u šťovíku.



Kvetení
[editovat | editovat zdroj]Jako kvetení je možné chápat dobu od otevření poupěte až do odkvětu, v této době dozrává pyl a vajíčka.
Otevírání květu
[editovat | editovat zdroj]Před otevřením květu rostlina vytvoří poupě, které do určité míry připomíná listové pupeny. Poupě je chráněno kališními lístky či vnějšími částmi okvětí. Otevírání poupěte patří mezi tropismy. Mohou se na něm podílet tyto mechanizmy:
- Vratná akumulace iontů. To bylo prokázáno pouze u hořce bezlodyžného (Gentiana acaulis).
- Apoptóza buněk v oblasti květních obalů.
- Ztráta vody během dne a její doplnění v noci. K tomu dochází např. u silenky Silene saxifraga, která rozkvétá na noc. Její korunní lístky se ve dne díky nedostatku vody svinou.
- Různá rychlost růstu vnějších a vnitřních květních obalů. U tulipánu vnější kruh okvětních lístků nejrychleji roste při teplotě o 10 °C nižší než vnitřní kruh, tulipán se tedy ráno otevře a večer uzavře.
Důležitým faktorem pro rozvinutí korunních lístků je zvětšení turgoru, které je obvykle způsobeno nárůstem koncentrace jednoduchých cukrů vznikajících ze složitých sacharidů (škrob např. v růžích, fruktany např. v denivkách). To je doprovázeno napnutím buněčných stěn. O vlivu rostlinných hormonů na otevírání květu jsou v literatuře protichůdné informace.
Otevírání květů spouštějí vnější faktory. U druhů rostlin otevírajících se na noc je hlavním spouštěčem večerní vlhkost. Na změny teplot reagují hlavně rostliny kvetoucí brzy zjara, např. sněženka (Galanthus nivalis) nebo šafrán (Crocus spp.). Dále mohou rostlin reagovat na světlo, jako např. sedmikráska (Bellis perennis). Otevírání a zavírání vytrvalých květů ve dne a v noci probíhá stejnými mechanizmy a je řízeno vnitřními biorytmy. O buněčné fyziologii a genetickém základu otevírání a zavírání květů se dosud mnoho neví.
Zavírání květů může probíhat díky rozdílné rychlosti růstu květních částí nebo vratné změny turgoru. V takovém případně je možné opakované otevírání a zavírání. Ztráta turgoru při odkvětu je nevratná.
Opylení
[editovat | editovat zdroj]Opylení je přenos pylu na samičí pohlavní orgány. U krytosemenných rostlin se jedná o přenos pylu na bliznu. Opylení a oplození jsou rozdílné děje. Květ může být opylen vlastním pylem (samosprašnost, autogamie) nebo pylem jiného jedince (cizosprašnost, allogamie).
Samosprašnost vede ke snižování genetické variability. U rostlin se vyvinuly různé způsoby, jak samoopylení zabránit:
- Prostorové oddělení prašníků a blizen (herkogamie). Např. u kapary trnité je pestík umístěn na vyvýšeném gynoforu, aby tyčinky nestály v cestě přilétajícímu hmyzu.

Herkogamie: u kapar se pestík nachází nad tyčinkami (zde kapara trnitá). - Prašníky a blizny mohou fungovat v jiném čase (dichogamie). Pokud se nejdříve uvolní pyl z prašných pouzder, jedná se o proterandrii; pokud jsou dříve funkční blizny, jedná se o proterogynii. Pokud se současně uvolňuje pyl a jsou funkční blizny, jde o homogamii.

Dichogamie a herkogamie mohou fungovat pouze v rámci jedné rostliny, nikoli v rámci různých rostlin. Proto existují další přizpůsobení:
- Vzájemná nekompatibilita: ta je zajištěna díky genetickým faktorům a zabraňuje samooplození. Někdy se vzájemná nekompatibilita projevuje i v morfologii, známým příkladem je různočnělečnost (heterostylie) u prvosenek (Primula).
Existují tři hlavní způsoby opylení: opylení větrem (anemogamie), vodou (hydrogamie) a prostřednictvím živočichů (zoogamie). Původní krytosemenné byly opylované primárně živočichy. Až druhotně se uzpůsobily opylování větrem či vodou. Mezi hlavní charakteristiky jednotlivých druhů opylování patří:
- Anemogamie: nenápadné květy; redukovaný počet orgánů; častá jedno- či dvoudomost; hustá, často nící květenství; nelepivý a hladký pyl; velké blizny; málo vajíček či jedno vajíčko v květu; nepřítomnost nektaru.
- Hydrogamie: nenápadné květy; častá jedno- či dvoudomost; výskyt provzdušňujících pletiv; smáčivý povrch pylových zrn; soudkovitá pylová zrna; velké blizny; málo vajíček či jedno vajíčko v květu.


- Zoogamie: U rostlin opylovaných živočichy jsou květy hlavně funkčními jednotkami. Rostlina může mít jeden květ (jako např. u tulipánu), nebo více květů uspořádaných v květenství, které jeden květ připomíná (pseudanthium, biologický květ). Tak tomu je u hvězdnicovitých (např. u sedmikrásky) nebo miříkovitých (např. mrkev). Zoogamní rostliny méně často vytvářejí více oddělených květů (např. u kosatce či u dřevin). Mezi hlavní znaky rostlin opylovaných živočichy patří: oboupohlavné květy nebo pseudanthia; výrazné zbarvení; silná vůně; pyl nebo nektar jako lákadlo pro opylovače, nebo šálení opylovačů; nerovný povrch pylových zrn, lepkavý pyl.
Oplození
[editovat | editovat zdroj]Pylové zrno se dostane na bliznu pestíku. Začne klíčit v pylovou láčku, která prochází zvláštním pletivem ve čnělce a dostane se až k vajíčkům v semeníku. V některých případech pylová láčka z určité blizny může prorůst jen k určité části semeníku.
Jakmile pylové láčky, resp. spermatozoidy z nich vzniklé dosáhnou vaječných buněk, může dojít k oplození. U krytosemenných dochází ke dvojímu oplození: splynutím spermatické buňky a vaječné buňky (oosféry) vzniká zygota, druhá spermatická buňka se spojí s jádrem zárodečného vaku a vytvoří tak triploidní živné pletivo, které vyživuje zárodek v semeni.
Po oplození se ze zygoty stane zárodek, z vajíčka se stane semeno a z květu (zejména semeníku) se stane plod.
Odkvět
[editovat | editovat zdroj]Rostliny na rozdíl od živočichů většinou dokážou vytvářet stále nové reprodukční orgány. Životnost jednotlivého květu je omezená, neboť květ spotřebovává množství zdrojů a blizny mohou být vstupní branou pro patogeny. Pokud by na rostliny vytrvávaly již opylené květy, vytvářela by se zbytečná konkurence ve spojitosti s opylovači.
Opylení proto vede k odkvětu (senescenci). Většina druhů rostlin dokáže zkrátit životnost jednotlivých květů. Opylení u mnoha rostlin spouští tvorbu hormonu ethylenu, který vyvolává stárnutí květních obalů. Některé druhy na ethylen nejsou citlivé, zde mechanizmus stárnutí květů není známý. Stárnou i další části květu: tyčinky a blizna se čnělkou, zatímco semeník se vyvíjí v plod. Před odkvětem se rostlina snaží vstřebat co nejvíce látek obsažených v květu, aby je mohla znovu využít – podobně jako se děje před opadem listů.[11]
Evoluce
[editovat | editovat zdroj]Evoluce květů krytosemenných není dostatečně vyjasněná, protože neexistují fosilní důkazy předchůdců „moderních“ květů. Také nejsou zcela jasné vztahy krytosemenných a dalších semenných rostlin. Existují dvě různé hypotézy o původu květů. Důvodem pro vznik oboupohlavného květu krytosemenných bylo dost možná opylování hmyzem (brouky), uzavření vajíček do pestíku mohlo být ochranou proti jejich konzumaci.
Euanthiová teorie Arbera a Parkina (1907)[12] předpokládá, že předkové krytosemenných rostlin měli oboupohlavné květy, květ je tedy jednoosým útvarem s postranně uspořádanými mikro- a megasporofyly. Tyčinky a pestíky by tedy vývojově odpovídaly listům. Tato teorie je založena na analýze fosilního rodu Caytonia, který je považován za možného předchůdce krytosemenných. Caytonia měla zpeřené mikro- a megasporofyly, které byly uspořádané v oboupohlavných útvarech. Na megasporofylech byly výstupky s větším množstvím vajíček po stranách. I mikrosporofyly byly zpeřené, každý mikrosporofyl nesl více skupin prašných váčků. Plodolisty krytosemenných mohly vzniknout redukcí rozvětvených megasporofylů, tyčinky možná vznikly díky redukci na nitku se dvěma prašnými pouzdry, kde v každém jsou dva prašné váčky.
Pseudanthiová teorie od Richarda Wettsteina praví, že květ krytosemenných vznikl z květenství jednopohlavných květů. Tyčinky a plodolisty by tak pocházely z oddělených výhonů. Základem této teorie je předpoklad, že krytosemenné rostliny vznikly z rostlin obalosemenných. Plodolisty tak měly vzniknout z listenů, které u obalosemenných chrání vajíčka. Tuto teorii nepodporují molekulární a morfologické analýzy.
Ačkoli se preferuje euanthiová teorie, původ tyčinek, plodolistů a druhého obalu vajíček je stále nejasný.
Teorie založené na molekulární biologii
[editovat | editovat zdroj]Nejstarší květy krytosemenných byly oboupohlavné a měly nerozlišené okvětí uspořádané do spirály.[2]
Mimo skupinu semenných rostlin nejsou rozmnožovací orgány stejného původu (nejsou homologní). U nahosemenných rostlin jednopohlavné šištice odpovídají skupinám genů B a C. Jejich exprese je podobná jako u krytosemenných: C se projevují ve všech reprodukčních orgánech, B v samčích šišticích. Na základě tohoto vznikly další hypotézy o původu oboupohlavných květů krytosemenných rostlin:
- Hypotéza „out of male“ od Theissena et al. (2002) tvrdí, že samčí šištice tvořily ve svých koncových částech samičí orgány díky potlačení exprese genů skupiny B. Podle hypotézy „out of female“ se samčí orgány vytvářely ve spodní části samičích šištic. Tyto modely předpokládají, že květní obaly vznikly až poté, co se vyvinulo oboupohlavné uspořádání.
- Hypotéza „mostly male“ vychází ze samčích květů, poprvé byla navržena Frohlichem a Parkerem (2000).[13] Tvrdí, že nahosemenné mají gen Leafy ve dvou kopiích. Gen Leafy se podílí na vzniku samčích květů, Needly na vzniku květů samičích. Needly se vyskytuje u všech nahosemenných kromě liánovců, ale ne u krytosemenných. Tato teorie tedy říká, že díky deaktivaci genu Needly se samčí šištice staly oboupohlavnými.[14]
V průběhu evoluce květu krytosemenných se musely odehrát čtyři klíčové události:
- Evoluce oboupohlavného květního stonku.
- Evoluce komprimovaného květního stonku a omezení růstu: Toho bylo dosaženo díky genům třídy C. Ty potlačují gen Wuchsel, který udržuje meristémy v činnosti.
- Evoluce květních obalů.
- Evoluce květních obalů vyšších dvouděložných rostlin rozdělených na kalich a korunu.
Soltis et al. (2007) uvažují o hypotéze, která vznik krytosemenných nezakládá na modelu ABC fungujícím u huseníčku, ale na modelu postupných přechodů jako např. u stulíku. Z toho by bylo možné odvozovat jak model ABC, tak jeho varianty (jako např. u tulipánu).[15]
Využití člověkem
[editovat | editovat zdroj]Květy se konzumují méně často než plody. Květenství květáku a artyčoku se využívají jako zelenina, z květenství konopí setého lze vytvořit kuřivo. Z jednotlivých květů či květenství lze vyrobit kupříkladu salát, nebo jimi lze zdobit jídla (violky, sedmikrásky, lichořeřišnice). Květy, poupata, části květů nebo květenství se využívají jako koření: pelyněk (Artemisia vulgaris subsp. vulgaris), levandule (Lavandula spp.), hřebíček (Syzygium aromaticum), šafrán (Crocus sativus) či chmel (Humulus lupulus).[16]
Větší roli hrají květy u kulturních rostlin, které se někdy pěstují i k řezu.
Bohyně červánků (Aurora/Eós) se zobrazovala s rozházenými květy odkazujícími na sluneční paprsky nového dne. Květy jsou také atributem bohyně Flory. Ze čtyř ročních období jsou květy nejvíce spojovány s jarem, ze základních smyslů s čichem. Květy jsou již od dob antiky symbolem pomíjivosti krásy a života. V malbách a kresbách zátiší se jako symbol pomíjivosti objevují zvadlé květy nebo opadané květní obaly. Alegorické postavy Logiky a Naděje jsou často vyobrazovány s květinami v náručí, neboť z květů se vyvíjejí plody.[17] Různé druhy rostlin mají vlastní symbolické významy, které se v průběhu staletí mění. Růže byla a je atributem Venuše, Dionýsa a Marie, je symbolem pro pozemskou i boží lásku, pro mladé ženy, ale i pro prostituci.[18]
V umění jsou květiny a květy součástí florálních motivů, kterými se často symbolizuje mládí či radost.
Odkazy
[editovat | editovat zdroj]Reference
[editovat | editovat zdroj]V tomto článku byl použit překlad textu z článku Blüte na německé Wikipedii.
- ↑ KVAČEK, Zlatko; KVAČEK, Jiří. Jak vznikly krytosemenné rostliny a jejich květy. Živa [online]. 2009 [cit. 2020-03-29]. Dostupné online.
- ↑ a b c d e f g THEISSEN, G.; MELZER, R. Molecular Mechanisms Underlying Origin and Diversification of the Angiosperm Flower. Annals of Botany. 2007-08-08, roč. 100, čís. 3, s. 603–619. Dostupné online [cit. 2020-03-27]. ISSN 0305-7364. DOI 10.1093/aob/mcm143. PMID 17670752. (anglicky)
- ↑ Lehrbuch der Botanik für Hochschulen. 35. Aufl. vyd. Heidelberg: Spektrum, Akad. Verl XIV, 1123 Seiten s. Dostupné online. ISBN 3-8274-1010-X, ISBN 978-3-8274-1010-8. OCLC 231972838
- ↑ Lexikon der Biologie. Heidelberg: Spektrum Akad. Verl CD-ROM s. Dostupné online. ISBN 3-8274-0341-3, ISBN 978-3-8274-0341-4. OCLC 174377864
- ↑ a b VINTER, Vladimír. Atlas anatomie cévnatých rostlin [online]. Olomouc: Katedra botaniky PřF UP, 2004 [cit. 2020-03-28]. Dostupné online.
- ↑ SLAVÍKOVÁ, ZDEŇKA, 1935-. Morfologie rostlin. 1. vyd. vyd. Praha: Karolinum 218 s. Dostupné online. ISBN 80-246-0327-6, ISBN 978-80-246-0327-8. OCLC 53273093
- ↑ RONSE DE CRAENE, LOUIS PHILIPPE. Floral diagrams an aid to understanding flower morphology and evolution. Cambridge: Cambridge University Press 1 Online-Ressource s. Dostupné online. ISBN 978-0-521-49346-8, ISBN 0-521-49346-3. OCLC 850992107
- ↑ JAEGER, Katja E.; GRAF, Alexander; WIGGE, Philip A. The control of flowering in time and space. Journal of Experimental Botany. 2006-10-01, roč. 57, čís. 13, s. 3415–3418. Dostupné online [cit. 2020-03-28]. ISSN 0022-0957. DOI 10.1093/jxb/erl159. (anglicky)
- ↑ CORBESIER, Laurent; COUPLAND, George. The quest for florigen: a review of recent progress. Journal of Experimental Botany. 2006-10-01, roč. 57, čís. 13, s. 3395–3403. Dostupné online [cit. 2020-03-28]. ISSN 0022-0957. DOI 10.1093/jxb/erl095. (anglicky)
- ↑ COEN, Enrico S.; MEYEROWITZ, Elliot M. The war of the whorls: genetic interactions controlling flower development. Nature. 1991-09, roč. 353, čís. 6339, s. 31–37. Dostupné online [cit. 2020-03-28]. ISSN 0028-0836. DOI 10.1038/353031a0. (anglicky)
- ↑ ROGERS, Hilary J. Programmed Cell Death in Floral Organs: How and Why do Flowers Die?. Annals of Botany. 2006-03-01, roč. 97, čís. 3, s. 309–315. Dostupné online [cit. 2020-03-28]. ISSN 1095-8290. DOI 10.1093/aob/mcj051. PMID 16394024. (anglicky)
- ↑ ARBER, E. A. Newell; PARKIN, John. On the Origin of Angiosperms.. Journal of the Linnean Society of London, Botany. 1907-07, roč. 38, čís. 263, s. 29–80. Dostupné online [cit. 2020-03-28]. DOI 10.1111/j.1095-8339.1907.tb01074.x. (anglicky)
- ↑ FROHLICH, Michael W.; PARKER, David S. The Mostly Male Theory of Flower Evolutionary Origins: From Genes to Fossils. Systematic Botany. 2000-04, roč. 25, čís. 2, s. 155. Dostupné online [cit. 2020-03-28]. DOI 10.2307/2666635.
- ↑ BATEMAN, R. M.; HILTON, J.; RUDALL, P. J. Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers. Journal of Experimental Botany. 2006-09-15, roč. 57, čís. 13, s. 3471–3503. Dostupné online [cit. 2020-03-28]. ISSN 0022-0957. DOI 10.1093/jxb/erl128. (anglicky)
- ↑ SOLTIS, Douglas E.; CHANDERBALI, André S.; KIM, Sangtae. The ABC Model and its Applicability to Basal Angiosperms. Annals of Botany. 2007-08-01, roč. 100, čís. 2, s. 155–163. Dostupné online [cit. 2020-03-28]. ISSN 1095-8290. DOI 10.1093/aob/mcm117. PMID 17616563. (anglicky)
- ↑ FRANKE, WOLFGANG. Nutzpflanzenkunde : nutzbare Gewächse der gemässigten Breiten, Subtropen und Tropen. 4., neubearb. u. erw. Aufl. vyd. Stuttgart: G. Thieme XI, 490 Seiten s. Dostupné online. ISBN 3-13-530404-3, ISBN 978-3-13-530404-5. OCLC 611494157
- ↑ Die Natur und ihre Symbole : Pflanzen, Tiere und Fabelwesen. Berlin: Parthas 382 s. Dostupné online. ISBN 3-936324-03-4, ISBN 978-3-936324-03-7. OCLC 66645437
- ↑ BEUCHERT, MARIANNE. Symbolik der Pflanzen. 1. Aufl. vyd. Frankfurt am Main: Insel-Verl 391 Seiten s. Dostupné online. ISBN 3-458-34694-5, ISBN 978-3-458-34694-4. OCLC 249217049
Související články
[editovat | editovat zdroj]Externí odkazy
[editovat | editovat zdroj] Obrázky, zvuky či videa k tématu květ na Wikimedia Commons
Obrázky, zvuky či videa k tématu květ na Wikimedia Commons  Téma Květ ve Wikicitátech
Téma Květ ve Wikicitátech Slovníkové heslo květ ve Wikislovníku
Slovníkové heslo květ ve Wikislovníku