
Cytokinin
Cytokininy jsou rostlinné hormony (fytohormony), které podporují dělení rostlinných buněk (cytokinezi), a tím u rostlin hrají důležitou roli v mnoha vývojových procesech. Nezbytné jsou pro podporu aktivního růstu a diferenciaci rostlinných buněk. Účastní se klíčení semen, vývoje kořene a prýtu (zejména vodivých pletiv), řídí stárnutí listů a velikost semen a odpověď na změny vnějšího prostředí a stres. Jejich funkce je často spojena s funkcí ostatních fytohormonů, zejména auxinu anebo ethylenu.
Historie
[editovat | editovat zdroj]Cytokininy byly objeveny rostlinným fyziologem švédského původu Folke K. Skoogem (1908–2001) a jeho spolupracovníky v 50. letech 20. století na univerzitě ve Wisconsinu. Látky s cytokininovou aktivitou byly nejprve nalezeny v kokosovém mléce a autoklávovaném sledím spermatu při hledání složek živného média nezbytných pro kultivaci rostlinných explantátů in vitro. První čistý cytokinin, kinetin, byl izolován v roce 1955 a jeho chemická struktura byla objasněna v roce 1956: jednalo se o 6-furfurylaminopurin.
Struktura
[editovat | editovat zdroj]Cytokinin je označení pro více chemických struktur s podobnou syntézou, degradací, funkcí etc.. Patří sem cis- i trans-zeatin, dihydrozeatin a izopentenyladenin. Mezi umělé analogy cytokininu, které rostliny syntetizovat neumí, ale pokud jsou do nich vpraveny uměle, fungují jako přirozené cytokininy, patří benzyladenin. Cytokininy jsou aktivní ve formě volných bází, reverzibilně je možné je inaktivovat konjugací s glukózou přes atom kyslíku či dusíku nebo s alaninem přes dusík. Konjugace je možná také s ribózou za vzniku ribosidů či ribotidů. Ireverzibilní inaktivace je možná odštěpením postranního řetězce cytokininoxidázou.
-
 Kinetin, první identifikovaný cytokinin
Kinetin, první identifikovaný cytokinin -
 Zeatin, isoprenoidní cytokinin
Zeatin, isoprenoidní cytokinin -
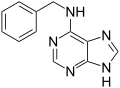 Benzyladenin, aromatický cytokinin
Benzyladenin, aromatický cytokinin



Biosyntéza a transport
[editovat | editovat zdroj]Biosyntéza aromatických cytokininů (benzyladenin, kinetin), není dosud známá. Oproti tomu enzymy zodpovědné za syntézu isoprenoidních cytokininů jsou známé od 80. let.
Primárním produktem biosyntézy jsou ribotidy vzniklé isoprenylací u rostlin ADP nebo ATP, u bakterií AMP.[1] Donorem postranního řetězce je dimethylallyl-pyrofosfát nebo (E)-hydroxymethylbutenyl-difosfát pro vznik isopentenyladeninu, respektive trans-zeatinu. Vzniklé ribotidy jsou aktivovány hydrolýzou enzymy rodiny LONELY GUY (podle fenotypu mutantní rýže s jediným klasem, na základě jehož byl tento gen a následně protein identifikován).[2] Jiný cytokinin, méně aktivní cis-zeatin, vzniká prenylací adenosinu v určitých molekulách tRNA a následnou hydrolýzou těchto molekul; ovšem fyziologická relevance této cesty stále ještě čeká na přímý biologický důkaz.
Původně se soudilo, že se tvoří převážně v kořenech, odkud jsou transportovány do nadzemní části. Ovšem studium exprese (výskytu) enzymů, které je syntetizují (tzv. isopentenyltransferáz) prokázalo, že k syntéze dochází prakticky v celé rostlině. Ukazuje se, že místem biosyntézy jsou pletiva, kde dochází ve vysoké míře k dělení buněk (tedy v převážné míře meristémy). Některé novější práce podávají důkazy, že cytokininy mohou být transportovány i z nadzemních částí do kořenů. Transportuje se xylémem (dle klasických schémat z kořene vzhůru) a dle některých poznatků rovněž i floémem (do kořenů). Na kratší vzdálenosti je přenášen transportéry rodiny PUP. Transportní forma jsou jejich deriváty ribosidy nebo ribotidy.
Degradace cytokininů je zprostředkovaná v zásadní míře proteiny z rodiny cytokinin oxidáz.
Přenos signálu
[editovat | editovat zdroj]Cytokininový receptor patřící mezi histidin kinázy je přítomen na membráně endoplazmatického retikula. Po vazbě cytokininu vytvoří dimer a tento protein se transautofosforyluje na histidinu (jinými slovy, právě zdimerizované podjednotky se fosforylují navzájem). Poté je fosfát přenesen na aspartát na témže proteinu. Z aspartátu se přenese na histidin proteinu HPT (histidine phosphotransfer, též z zvané AHP), který je transportován do jádra, kde ovlivní genovou expresi pomocí některých z proteinů ARR.
Fyziologické funkce
[editovat | editovat zdroj]- stimulace dělení buněk
- inhibice apikální dominance (viz auxin) a tím stimulace větvení
- inhibice tvorby protoxylému v kořeni a také tvorby postranních kořenů
- regulace aktivity apikálních meristémů
- tvorba druhotných pletiv a stimulace druhotného tloustnutí v kořeni i stonku
- oddálení senescence
- stimulace růstu kořenů a klíčení
- stimulace fotosyntézy[3]
Reference
[editovat | editovat zdroj]- ↑ FRÉBORT, Ivo; KOWALSKA, Marta; HLUSKA, Tomáš. Evolution of cytokinin biosynthesis and degradation. Journal of Experimental Botany. 2011-05-01, roč. 62, čís. 8, s. 2431–2452. PMID: 21321050. Dostupné online [cit. 2016-04-27]. ISSN 0022-0957. DOI 10.1093/jxb/err004. PMID 21321050. (anglicky)
- ↑ KURAKAWA, Takashi; UEDA, Nanae; MAEKAWA, Masahiko. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature. Roč. 445, čís. 7128, s. 652–655. Dostupné online. DOI 10.1038/nature05504.
- ↑ Mohr, Schopfer. Plant Physiology. [s.l.]: Springer-Verlag Berlin, 1995. Dostupné online. (anglicky)
Externí odkazy
[editovat | editovat zdroj] Obrázky, zvuky či videa k tématu cytokinin na Wikimedia Commons
Obrázky, zvuky či videa k tématu cytokinin na Wikimedia Commons - A Companion to Plant Physiology: Cytokinins: Regulators of Cell Division Archivováno 11. 3. 2011 na Wayback Machine. (anglicky)
- Plant Physiology (Biology 327): Plant Hormones - Cytokinins (anglicky)
