
Pískovník rybářský
 | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | kroužkovci (Annelida) |
| Podkmen | bezopaskovci (Aclitellata) |
| Třída | mnohoštětinatci (Polychaeta) |
| Řád | Capitellida |
| Čeleď | Arenicolidae |
| Rod | pískovník (Arenicola) |
| Binomické jméno | |
| Arenicola marina (Linnaeus, 1758) | |
| Synonyma | |
Lumbricus marinus Linnaeus, 1758 | |
| Některá data mohou pocházet z datové položky. | |
Pískovník rybářský (Arenicola marina) je kroužkovec, jehož tělo se skládá z řady po sobě jdoucích kroužků, tzv. segmentů nebo metamerů. U pískovníka jsou tyto segmenty rozděleny na 2 až 5 povrchových prstenců. Pískovník patří do třídy mnohoštětinatců (Polychaeta, od poly = několik a chaete = štětiny): segmenty nejrobustnější části jsou vybaveny parapodii opatřenými více či méně tuhými pohyblivými štětinami, které umožňují živočichovi opřít se o podklad, aby se znehybnil nebo se pohyboval. V dospělosti dosahuje průměrné délky kolem patnácti centimetrů, maximálně však kolem třiceti centimetrů.[1] Slovo Arenicola je složeno z latinského arena = písek + colere = obývat,[2] slovo marina = mořská.[3]
Místní pojmenování
[editovat | editovat zdroj]Francouzsky je nazýván jako « arénicole », či « ver de vase » nebo « ver noir » a je dobře znám francouzským rybářům, kteří jej nazývají několika dalšími místními názvy, jako je chique, buzuc, buzin atd. Existuje i název specifický pro západní Normandii (Calvados, Manche) sando[n] nebo sandron ze staroseverského *sandormr, od sandr „písek“ a ormr „had“, srov. dánské sandorm, stejný význam.
Morfologie
[editovat | editovat zdroj]Tělo je přibližně válcovité, v přední části silně zduřelé a v zadní části užší.[4] Poněkud konvenčně jej lze rozdělit na čtyři části: hlava, hruď, břicho a ocas.
Wells[5] spojuje hruď a břicho pod názvem „trup“, který tvoří 19 segmentů vybavených parapodii („setigery“ = nesoucí sety), které Wells čísluje i–xix.
V české literatuře se oddíly objevují např. jako příď (přída) a záď, případně je tělo členěno na hlavu, střední a ocasní část.[6]
Hlava
[editovat | editovat zdroj]Hlava se skládá z prostomia, peristomia a segmentu bez parapodií a štětin.
Prostomium, čelní lalok, je malý trojúhelníkový hřbetní výběžek, jehož základna je otočena dopředu a je umístěna za ústy. Jeho povrch je rozdělen na tři laloky dvěma rýhami uspořádanými do tvaru písmene „Y“ a neobsahuje žádný zvláštní orgán. Může být vtaženo do vaku, nuchálního vaku, a proto není okamžitě viditelné.
Peristomium, známé také jako bukální segment, obsahuje ústa vybavená okrouhlou exserózní trubicí – hltanová trubice, která se může vychlipovat vně a je pokryta četnými papilami. Tento segment odpovídá prvním dvěma prstencům přední části, ne vždy jasně odděleným.
Další dva prstence představují segment bez výrůstků („bez štětin“).
Hruď
[editovat | editovat zdroj]Hruď je tvořena šesti segmenty opatřenými štětinami („setigery“). První segment má dva prstence, druhý tři, třetí čtyři a pátý a šestý pět. Parapody jsou umístěny na předposledním prstenci každého segmentu, ten je výraznější než ostatní. Každý parapod se skládá z hřbetního laloku neboli notopodu (od noto = hřbet a pode = noha) opatřeného dlouhými ostnitými sety a břišního laloku neboli neuropodu (umístěného na straně nervového provazce, který je ventrální). Neuropod představuje v případě pískovníka nízký lalok v laterálně ventrálním směru, který je vybaven krátkými, esovitými sety známými jako „uncinis“ (= háčky). Dle Wellse segmenty i–vi.
Břicho
[editovat | editovat zdroj]Břicho je tvořeno následujícími 13 segmenty, které jsou podobné posledním segmentům hrudníku, ale jsou vybaveny rozvětvenými, kontraktilními, červenými žábrami umístěnými v blízkosti hřbetní části parapodia. Oddíl s keříčkovitými žábrami je zbarven znatelně červeně. Dle Wellse segmenty vii–xix.
Ocas
[editovat | editovat zdroj]Ocas je zřetelně užší než předchozí oddíl a je tvořen neurčitým počtem segmentů bez parapodií a žaber, z nichž každý je reprezentován jedním prstencem. Segmenty jsou na bázi ocasního oddílu velmi krátké a na konci směrem k řitnímu otvoru jsou prodloužené. Pokožka pískovníků vylučuje šedozelenou fluoreskující látku, která je poněkud dráždivá a při manipulaci zbarvuje prsty.
Orgánové soustavy
[editovat | editovat zdroj]Orgánové soustavy viz Tixier[7] a Wells[5].
- Trávicí ústrojí: zahrnuje rypáček, následuje jícen se dvěma divertikly (vychlípeniny), jícnové žlázy, pak přichází vypouklý žaludek, pokrytý žlutou tkání, chloragogenní tkáň (tkáň, která hromadí odpadní látky, ale pravděpodobně slouží i jako zásobárna různých látek) a nakonec dlouhé střevo končící řití.
- Oběhová soustava: Krev je obsažena ve složitém systému uzavřených cév; je zbarvena červeně erytrokruorinem rozpuštěným v plazmě a uváděna do pohybu dvěma laterálními srdci (každé má 1 předsíň a 1 komoru) a cévami se stahujícími se stěnami.
- Vylučovací a pohlavní ústrojí: 6 párů nefridií (3 hrudní + 3 břišní, dle Wellse segmenty iv–ix), které se skládají z nálevky s vířivým epitelem (nefrostoma), nefridiového váčku a stahovatelného měchýřku, který se otevírá do podlouhlého póru (nefridiopóru) za horní úrovní neuropodů. Gonády se nacházejí na zadním konci nálevky posledních 5 nefridií, které zajišťují evakuaci pohlavních produktů, a jsou často skryty váčkem.
- Nervová soustava: Mozková ganglia se špatně vymezenými obrysy jsou přitisknuta ke „stropu“ prostomia (čelní lalok), následuje perioezofageální límec (= vazivo) a pak podélný nervový provazec bez ganglií, jehož přítomnost je navenek prozrazena bělavou linií na dně rýhy na ventrální straně pískovníka. V bukálním segmentu jsou dvě žluté statocysty. Tyto smyslové orgány umožňují pískovníkovi poznat svou polohu v prostoru (zda je ve vodorovné nebo svislé poloze, zda je jeho hlava otočena nahoru nebo dolů atd.).
Biologie
[editovat | editovat zdroj]Život v sedimentech
[editovat | editovat zdroj]
Pískovník se vyskytuje v hloubce 10 až 20 cm, nebo dokonce 40 cm, v písčitých nebo písčito-bahnitých sedimentech. Obecně se nevyskytuje v sedimentech s průměrnou velikostí částic menší než 80 μm a jeho hojnost klesá, když tato velikost přesáhne 200 μm.[p 1] Červ vytváří chodbičku ve tvaru písmene „U“, která se skládá ze dvou částí.[8]
Chodbička
[editovat | editovat zdroj]Chodbička je trubice přibližně ve tvaru písmene „L“ nebo „J“ (podle strany pozorovatele), která zabírá vodorovnou a jednu svislou větev písmene „U“. Je vystlána světlým sedimentem (šedým nebo rezavým, což ukazuje na jeho oxidovaný stav) konsolidovaným hlenem vylučovaným živočichem. Horní část vertikální větve má užší část, kaudální „komín“, do kterého proniká pouze ocas červa zejména při defekaci. Exkrementy tvořené převážně sedimentem vytvářejí neúhledné provazovitě stočené tvary nebo kužel více či méně vysoko nad okolním povrchem. Pískovník obvykle zabírá nejširší část chodbičky, tj. horizontální větev a spodní část vertikální větve.
Přední část
[editovat | editovat zdroj]Přední část představuje především druhou vzestupnou větev. Její tvar a struktura jsou velmi variabilní v závislosti na charakteru sedimentu a činnosti pískovníka. Jedná se o kuželovitý prostor s nálevkovitou prohlubní na povrchu, nebo dokonce o otvor vedoucí do chodbičky, který je však zpravidla vyplněn sedimentem, jenž byl silně přepracován živočichem, který sem sporadicky vniká, a proudem vody, který zde udržuje.
V tomto systému způsobuje pískovník dva proudy v opačných směrech: Proud vody je způsobený vlnami dilatace a kontrakce, které probíhají od zadní části těla k přední, a pohání vodu od ocasu k hlavě, přičemž prochází hlavně hřbetní stranou. Vytvořený proud činí 1,2 až 7,2 ml za minutu u živočicha o hmotnosti 17,2 g[9] a 1,5 ml za minutu u „standardního“ pískovníka (suchá hmotnost = 0,5 g) se 7 kontrakcemi za minutu při 15 °C.[10]
Kromě toho tento proud vody prosakuje sedimentem v přední části systému a mění jeho složení a stabilitu (vzniklý tlak dosahuje alespoň dočasně 20 cm vody).[10] Každý zavlažovací cyklus se skládá ze tří fází: a) zpětný posun těla, b) čerpání a posun těla vpřed, c) zpětné čerpání. Fáze b) je nejdůležitější.[11]
Cirkulace sedimentu je způsobená tím, že živočich polyká písek (částice < 2 mm) a bahno na bázi přední části a vyvrhuje je nad zadním ústím.
Pískovník je jedním z mořských psamivorních organismů (z psammivor, doslova „pískožrout“, odvozeno ze starořeckého psámmos a latinského vore). Červ se živí organickým detritem i mikroflórou a mikrofaunou obsaženou v sedimentu. Některé z těchto součástí pocházejí z usazenin, které se vytvářejí v povrchové prohlubni nad přední větví (odtud pochází termín deposivor – „usazeninožravec“, který je pískovníkovi přisuzován).[9] Byla vyslovena domněnka (Krüger, 1959), že posteroanteriorní vodní proud může obohacovat sediment o organické prvky, které s sebou nese, a přispívat tak k potravě pískovníků. Tato hypotéza se nezdá být platná.[10]
Tyto činnosti probíhají, když je biotop živočicha ponořen. Cykly „větrání“ v chodbičce trvají 10 až 15 minut[9] a jsou odděleny obdobími krmení (trvajícími přibližně 7 minut)[12] nebo odpočinku. Tyto cykly jsou řízeny nervovým systémem: kardiostimulátor v břišním řetězci pro ventilaci, další ve stěně jícnu pro krmení, jsou automatické a zřejmě nezávislé na vnějších podmínkách.[12]
Při malé vodě se v závislosti na odtokových podmínkách sedimentu může horizontální chodbička vyprázdnit, v takovém případě červ stojí ve vertikální zadní části a dýchá kyslík ze vzduchu v atmosféře nasycené vlhkostí, případně přitahuje vzduchové bubliny k žábrám.[8] Zůstane-li chodbička ve vodě, může živočich vyčerpat rozpuštěný kyslík a být vystaven prostředí s vysokou koncentrací sirovodíku (H2S, HS−), načež přejde do anaerobního metabolismu; při opětovném ponoření oxiduje sirovodík nahromaděný v coelomové tekutině na thiosíran. Z tohoto stresu se zotaví za méně než hodinu.[13]
Rozmnožování
[editovat | editovat zdroj]Pohlaví jsou oddělená a nelze je navenek rozpoznat. V průměru připadá 3,75 samic na 1 samce.[14][15] Z gonád, které jsou velmi malé, se uvolňují pohlavní buňky do célomu, kde se dále vyvíjejí. Oocyty jsou od sebe oddělené a snadno rozpoznatelné. Spermie (vlastně spermatocyty) jsou sestaveny do kulatých útvarů se vzhledem ostružiníku („morula“), které se před uvolněním rozpadají pod vlivem „faktoru zrání“ vylučovaného prostomiem.[16]
Rozmnožování vrcholí na podzim (říjen–listopad). Gamety se uvolňují současně, během krátkého období 4–5 dnů během malých vod skočného dmutí. Tato synchronizace je způsobena chemickými signály (feromony), které uvolňují rozmnožující se jedinci.[17] Oocyty zůstávají v chodbičkách, spermie se rozprostřou na povrchu sedimentu a jsou proudem vody, který vytvářejí samice při přílivu moře, zaneseny do chodbičky, kde dochází k oplození.
Larvy se vyvíjejí v samičí chodbičce.
Postlarvy se plazí v povrchové vrstvě sedimentu a mohou být unášeny proudy, ale nevedou planktonní život.
Mladí jedinci se usazují v jiných teritoriích, než která obývají starší jedinci.[18] Pohlavní dospělosti mohou dosáhnout po roce, zpravidla ve druhém roce života. Dožívají se nejméně 5–6 let. Podle Newella (1948) dosahuje úmrtnost dospělců po rozmnožení 40 %, jiní autoři uvádějí průměrnou roční úmrtnost 22 %.
Význam
[editovat | editovat zdroj]
Populace pískovníků mohou dosahovat hustoty 100–150 jedinců na metr čtvereční a tvoří 30 % biomasy běžné pláže.[19]
Jejich hrabání, čerpání a krmení může radikálně přepracovat sediment (jev známý jako bioturbace), změnit jeho strukturu (jemné částice < 2 mm se dostávají na povrch, zatímco hrubá frakce zůstává v hloubce) a také cirkulaci a fyzikálně-chemické vlastnosti vody, kterou obsahuje (zejména koncentraci kyslíku).
Tento druh proto může zanechat velmi výraznou stopu na pobřeží, které obývá.
Je důležitým zdrojem potravy pro platýse a několik druhů pobřežních ptáků. Predátorovi se zpravidla podaří ukořistit pouze koncovou část ocasu. Ocas získá svou původní délku prodloužením zbývajících segmentů. Tyto amputace se dějí bez většího poškození pískovníků, ačkoli při jejich provádění jednou týdně se růst zastaví.[20]
Pískovníci jsou velmi vyhledávanou nástrahou k lovu mořských ryb a jsou předmětem prosperujícího obchodu. Sbírají se vyrytím či vykopáním sedimentu, vytažením živočicha pomocí čerpadla (v písku) nebo (v Nizozemsku) pomocí speciálně vybavených člunů. Bylo spočítáno, že v Nizozemsku se ročně vytěží 29 až 36 milionů pískovníků.[21] Rozsáhlá těžba (několik stovek metrů čtverečních) může mít znepokojivé účinky na faunu, a dokonce i na flóru dotčeného pobřeží, i když se populace pískovníků během několika měsíců obnoví díky migraci jedinců (zejména mladých jedinců) ze sousedních oblastí.
Pískovník se používá v biotestech zkoumajících účinky toxinů v sedimentech a v různých studiích týkajících se procesů jako je dýchání. Jeho hemoglobin byl navržen jako náhrada lidské krve pro transfuze.[22]
Příbuzné druhy
[editovat | editovat zdroj]Z pěti druhů rodu Arenicola, které uvádí Fauvel, jsou v současnosti do rodu Arenicolides zařazeny druhy bez ocasu (A. ecaudata a A. grubii).[23] V evropských vodách byl od druhu A. marina oddělen nový druh Arenicola defodiens (uvádí ho v poznámce Fauvel).[24] Dosahuje větších rozměrů než A. marina, vyskytuje se na exponovanějších plážích a na nižší úrovni pobřeží.[25] Zdá se, že A. defodiens nebyl zaznamenán na pobřeží kontinentální Evropy a není uveden v seznamu druhů vyskytujících se v Lamanšském průlivu.[26]
Současné nebo potenciální lékařské využití
[editovat | editovat zdroj]Molekula, kterou tento červ používá k vázání kyslíku v krvi, je 250krát menší než lidská červená krvinka, ale 40krát více okysličuje, což má několik lékařských využití.[27] Mohla by znamenat revoluci v používání práškové krve, protože ji lze sušit mrazem, na rozdíl od současných roztoků (vyvinutých z lidských nebo hovězích buněk), které mají četné vedlejší účinky.
Společnost Hemarina se sídlem v Morlaix vyvinula a uvedla na trh univerzální nosič kyslíku, známý jako M101, založený na pískovníkově hemoglobinu.[28]
Galerie
[editovat | editovat zdroj]-
 Arenicola marina. Různé části těla.
Arenicola marina. Různé části těla. -
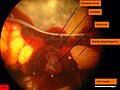 Arenicola marina. Pohled na pravé srdce. Komora je stažená a téměř prázdná. Žluté tečky jsou oocyty.
Arenicola marina. Pohled na pravé srdce. Komora je stažená a téměř prázdná. Žluté tečky jsou oocyty. -
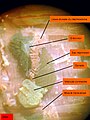 Arenicola marina. Pohled na nefridie a gonádu. Příčné svaly, které tyto orgány částečně zakrývají, byly rozříznuty.
Arenicola marina. Pohled na nefridie a gonádu. Příčné svaly, které tyto orgány částečně zakrývají, byly rozříznuty. -
 Arenicola marina. Nefrostoma při pohledu z ventrální strany a gonáda.
Arenicola marina. Nefrostoma při pohledu z ventrální strany a gonáda. -
 Arenicola marina. Nervový systém. Řez byl proveden na pravé straně hlavy, hřbetní část je přeložena doleva, prostomium je vidět z ventrální strany. Svalové svazky kryjí nervové elementy. Nad koncem abdukční linie je vidět „X“ tvořené setkáním dvou pojivových tkání a odchodem dvou velkých bílých provazců nervových vláken.
Arenicola marina. Nervový systém. Řez byl proveden na pravé straně hlavy, hřbetní část je přeložena doleva, prostomium je vidět z ventrální strany. Svalové svazky kryjí nervové elementy. Nad koncem abdukční linie je vidět „X“ tvořené setkáním dvou pojivových tkání a odchodem dvou velkých bílých provazců nervových vláken. -
 Arenicola marina. Čerstvý preparát statocysty pod mikroskopem.
Arenicola marina. Čerstvý preparát statocysty pod mikroskopem. -
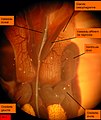 Arenicola marina. Hřbetní pohled na obě srdce. Obě síně nekomunikují s hřbetní cévou, ke které jsou připojeny. Chloragénní tkáň dole. Rozptýlené žluté tečky jsou oocyty, volně v coelomové tekutině.
Arenicola marina. Hřbetní pohled na obě srdce. Obě síně nekomunikují s hřbetní cévou, ke které jsou připojeny. Chloragénní tkáň dole. Rozptýlené žluté tečky jsou oocyty, volně v coelomové tekutině. -
 Arenicola marina. Boční pohled na přední břišní metamerium.
Arenicola marina. Boční pohled na přední břišní metamerium. -
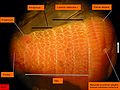 Arenicola marina. Boční pohled na hlavu.
Arenicola marina. Boční pohled na hlavu.









Odkazy
[editovat | editovat zdroj]Poznámky
[editovat | editovat zdroj]- ↑ Frakce jemný písek je definována průměrem částic 50 až 250 μm.
Reference
[editovat | editovat zdroj]V tomto článku byl použit překlad textu z článku Arenicola marina na francouzské Wikipedii.
- ↑ FOURRIER, Guillaume. La pêche à pied: dans votre poche. [s.l.]: [s.n.], 2014. 96 s. Dostupné online. ISBN 978-2-85725-916-9. S. 102. (francouzsky).
- ↑ POLÍVKA, František. Etymologický slovníček latinského názvosloví přírodopisného. Praha: Fr. A. Urbánek, 1883. 136 s. Dostupné online. S. 18.
- ↑ POLÍVKA, František. Etymologický slovníček latinského názvosloví přírodopisného. Praha: Fr. A. Urbánek, 1883. 136 s. Dostupné online. S. 77.
- ↑ Fauvel, P. 1927. Faune de France 16. Polychètes errantes. 494 p Téléchargeable sur :www.faunedefrance.org
- ↑ a b WELLS, George Philip. The anatomy of the body wall and appendages in Arenicola marina L., Arenicola claparedii Levinsen and Arenicola ecaudata Johnston. Journal of the Marine Biological Association of the United Kingdom. Cambridge: Marine Biological Association of the United Kingdom, 1950, Volume 29, Issue 1, s. 1–44. Dostupné online. ISSN 0025-3154. DOI 10.1017/S0025315400056174. (anglicky)
- ↑ REICHHOLF, Josef; JANKE, Klaus; KREMER, Bruno P. Moře a pobřeží: Ekologie mořských životních prostředí Evropy. Překlad Jiří Čihař. Praha: Ikar, 1999. ISBN 80-7202-409-4. S. 136.
- ↑ Tixier, A. & Gaillard, J. M. 1957. Anatomie animale et dissection. Vigot Frères éditeurs. 375 p.
- ↑ a b Wells, G. P., 1945. The mode of life of Arenicola marina L. Jour. Mar. Biol. Ass. U. K., 26, 170-207. (anglicky)
- ↑ a b c Toulmond, A. & Dejours, P. 1994. Energetics of the ventilatory piston pump of the lugworm, a deposit–feeding polychaete living in a burrow. Biological Bulletin, 186 : 213-220.
- ↑ a b c Riisgård, H. U., Berntsen, I & Tar, B. 1996. The lugworm (Arenicola marina) pump :characteristics, modelling and energy cost. Mar.Ecol. Prog.ser. 138 : 149-156.
- ↑ Wells, G. P., 1949a. Respiratory movements of Arenicola marina L.: intermittent irrigation of the tube, and intermittent aerial respiration., Jour. mar. biol. Ass. U. K., 28, 447-478.
- ↑ a b Wells G. P. 1949b. The behaviour of Arenicola marina in sand, and the role of spontaneuos activity cycles. Jour. Mar. biol Ass, UK 29:1-44.
- ↑ Völkel, S. Hauschild, K & Grieshaber, M. K. 1995. Sulfide stress and tolerance in the lugworm Arenicola marina during low tide. Mar. Ecol. Prog. Ser. 122 : 205-215.
- ↑ Newell, G. E., 1948. A contribution to our knowledge of the life history of Arenicola marina L.., Jour mar. biol. Ass. U.K. 28, 554-580.
- ↑ Howie, D. I. D., 1959. The spawning of Arenicola marina (L.). I. The breeding season. Jour. mar. biol. Ass. U.K. 38, 395-406.
- ↑ Pacey, A. A. & Bentley, M.G. 1992. The fatty acid 8,11,14-eicosatrienoic acid induces spawning in the male lugworm Arenicola marina. Jour.exp. Biol. 173 : 165-179.
- ↑ Hardege, J. D. & Bentley M. G. 1997. Spawning synchrony in Arenicola marina: evidence for sex pheromonal control. Proceedings:Biological Sciences, 264 : 1041-1047.
- ↑ Hardege, J. D., Bentley, M. G. & L. Snape. 1998. Sediment selection by juvenile Arenicola marina. Mar. Ecol. Prog.Ser. 166 : 187-195
- ↑ Arenicola marina : http://zipcodezoo.com/Animals/A/Arenicola_marina/
- ↑ Bergman, M. J. N., van der Veer, H. W. & Karczmarski, L. 1988. Impact of tail-nipping on mortality, growth and reproduction of Arenicola marina. Neth. J. Sea Res. 22: 83-90 www.vliz.be/vmdcdata/imis2/imis.php?module=ref&refid=15830
- ↑ McLusky, D. S. & Anderson, F. E. 1983. Distribution and population recovery of Arenicola marina and other benthic fauna after bait digging. Mar. Ecol.prog.ser. 11 : 173-179.
- ↑ Des chercheurs français inventent le sang de mer www.transfert.net/Des-chercheurs-français-inventent 27k-
- ↑ Fauchald, K. 1977. The polychaete worms. Definitions and Keys to the Orders, Families and Genera Natural History Museum of Los Angeles County, Science Series 28, 188p.
- ↑ Cadman, P. S. & Nelson-Smith, A. 1993. A new species of lugworm : Arenicola defodiens sp.nov. Jour. mar. biol. Ass.U.K. 73: 213-223
- ↑ Everson, C. Two species of lugworm. Dostupné online.
- ↑ Dauvin, J. C., Dewarumez, J. M. & Gentil, F. 2003. Liste actualisée des espèces d’Annélides Polychètes présentes en Manche. Cah. Bio. Mar. 44 : 67-95.
- ↑ Le Pape Fiona, a kol. (2015) Advancement in recombinant protein production using a marine oxygen carrier to enhance oxygen transfer in a CHO-S cell line. Artificial Cells, Nanomedicine, and Biotechnology. 43(3). doi: 10.3109/21691401.2015.1029632
- ↑ Anaximandre. HEMOXYCarrier - Français - Transporteur doxygène universel. www.hemarina.com [online]. [cit. 2020-03-09]. Dostupné v archivu pořízeném z originálu dne 2020-04-18.
Literatura
[editovat | editovat zdroj]- RULÍK, Martin. K biologii pískovníka rybářského. Živa. Academia, 1994, roč. 42, čís. 4, s. 174–175. ISSN 0044-4812.
Externí odkazy
[editovat | editovat zdroj] Obrázky, zvuky či videa k tématu pískovník rybářský na Wikimedia Commons
Obrázky, zvuky či videa k tématu pískovník rybářský na Wikimedia Commons