
Folikulární pomocné T lymfocyty
Folikulární pomocné T lymfocyty (zkratka TFH z angl. follicular helper T lymphocytes) patří do skupiny pomocných TH lymfocytů exprimujících povrchový znak koreceptor CD4 (též CD4+ T lymfocyty). Existence této lymfocytární populace byla poprvé reportována na přelomu tisíciletí v lidských krčních mandlích[1]. Označení těchto buněk jako „folikulární“ odkazuje na jejich lokalizaci především ve folikulech sekundárních lymfatických orgánů (uzliny, slezina). Tato lokalizace TFH lymfocytů dále napovídá jejich funkci v imunitním systému.[2][3]
TFH lymfocyty jsou důležitou složkou účastnící se adaptivní imunitní odpovědi a také nastavení imunitní paměti. Na těchto základních imunitních mechanismech se TFH lymfocyty podílejí zejména svou spoluprací s B lymfocyty na tvorbě specifické protilátkové odpovědi během reakcí germinálního centra (GC). Tyto interakce poté vedou ke klonální expanzi antigen-specifických B lymfocytů, jejich diferenciaci v protilátky produkující plazmatické buňky a také v dlouho žijící paměťové B buňky.[2][4]
Funkce TFH[editovat | editovat zdroj]
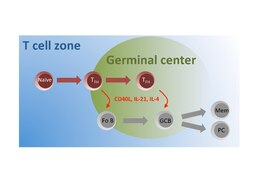
TFH lymfocyty jsou jednou z klíčových buněčných populací podílejících se na imunitních dějích v sekundárních lymfatických orgánech. TFH buňky jsou nutné pro správné vytvoření a chod GC v B buněčné zóně lymfoidních folikulů, na čemž kromě B lymfocytů spolupracují i s dalším buněčným typem který se zde nachází tzv. folikulární dendritické buňky (FDB, angl. FDC). Germinální centra jsou právě ta místa ve kterých dochází k zásadním procesům B buněčné imunitní odpovědi – afinitní maturaci B lymfocytů, která zahrnuje somatickou hypermutaci (SHM) B buněčného receptoru (BCR) a následnou selekci. Dále zde také dochází k izotypovému přesmyku (angl. class switch recombination (CSR)) a následně k produkci konkrétního izotypu protilátek. A v neposlední řadě zde dochází k diferenciaci v plazmatické a paměťové buňky.[5][6]
Na těchto procesech se TFH lymfocyty podílejí přímým buněčným kontaktem s B lymfocyty pomocí párů adhezivních a signalizačních molekul, jako jsou TCR a p:MHC-II, ICOS a ICOS-L, PD1 a PD1L a další. TFH lymfocyty zprostředkovávají kostimulační signál přes interakci molekul CD40L (na povrchu TFH lymfocytu) a CD40 (na povrchu B lymfocytu). V neposlední řadě jsou TFH lymfocyty zdrojem cytokinů, zejména IL-21 a IL-4, které jsou důležité pro proliferaci B lymfocytů. Spektrum cytokinů produkované TFH lymfocyty ovlivňuje také izotypový přesmyk, a tudíž i výsledný izotyp produkovaných protilátek (u člověka z IgD/IgM na IgG1, IgG2, IgG3, IgG4, IgA, IgE). Tyto interakce jsou vzájemné a B lymfocyty v kontaktu s TFH lymfocyty u nich podporují expresi kostimulačních molekul, produkci cytokinů a setrvání v GC.[7][8]
Diferenciace TFH[editovat | editovat zdroj]
Populace TFH lymfocytů je definována expresí transkripčního faktoru Bcl6, který je hlavním regulátorem jejich diferenciace a ustanovuje TFH buněčný program. Další znak, kterým je tato populace charakterizována je chemokinový receptor CXCR5, podle kterého byla tato populace poprvé definována a který TFH lymfocytům umožňuje lokalizaci do B buněčných folikulů. Typická je také exprese molekul PD1, ICOS a IL-21.[9][10]
Diferenciace TFH lymfocytů je úzce spojena se sekundárními lymfatickými orgány a průchodem těchto buněk přes T buněčnou a B buněčnou zónu. Zde k diferenciaci a dozrávání funkčních TFH lymfocytů dochází v několika krocích. V T buněčné zóně dochází ke stimulaci TCR nezkušených TN/TH0 buněk p:MHC-II komplexem na povrchu dendritických buněk, společně s dalšími kostimulačními a cytokinovými signály. Tyto interakce vedou k aktivaci exprese Bcl6 a diferenciaci TN/TH0 buněk do TFH linie. Takto diferencované TFH lymfocyty exprimují CXCR5 a zároveň snižují expresi CCR7 a migrují do B buněčné zóny, kde dochází k formování a reakcím GC. Interakce TFH lymfocytů v GC posílí expresi CXCR5, ICOS, PD1, sekreci cytokinů a takto aktivované TFH lymfocyty spolupracují s B lymfocyty na imunitní odpovědi.[8][11]
TFH a jejich role v patologiích[editovat | editovat zdroj]
Primární imunodeficience[editovat | editovat zdroj]
Jsou reportována onemocnění spojená s dysregulovanou nebo deficitní funkcí TFH lymfocytů. Pokud jsou TFH lymfocyty nefunkční dochází k nedostatečné či porušené produkci protilátek, což může vést k imunodeficitním stavům a přecitlivělosti k infekcím způsobenými zejména extracelulárními patogeny. Nejedná se pouze o deficity v samotné TFH buněčné populaci, ale také o deficity klíčových molekul nutných pro jejich diferenciaci a funkci. Pacienti s deficity v genech pro molekuly CD40L či ICOS trpí sníženým počtem TFH lymfocytů a defekty v produkci protilátek vedoucím až k hyper IgM syndromům či běžné variabilní imunodeficienci (angl. common variable immunodeficiency (CVID)).[9][12]
Získané imunodeficience[editovat | editovat zdroj]
Virus HIV cílí na pomocné TH lymfocyty jejichž součástí jsou i TFH lymfocyty, ty navíc mohou sloužit jako rezervoár této virové infekce. Výsledkem infekce virem HIV je pak kromě buněčných imunodeficitů a syndromu AIDS také neschopnost těchto pacientů vytvořit robustní specifickou protilátkovou odpověď proti patogenům.[12] Jelikož TFH lymfocyty jsou důležité i pro diferenciaci dlouho žijících paměťových B buněk, jejich nedostatečná funkce může také snížit nastavení imunitní paměti a účinnost vakcinace cílící na tento imunitní mechanismus, což bylo pozorováno u HIV pozitivních pacientů při očkování proti chřipce.[5][13]
Autoimunitní onemocnění[editovat | editovat zdroj]
U pacientů s některými autoimunitními onemocněními, jako je systémový lupus erythematodes (SLE) či revmatoidní artritida (RA) byly naměřeny zvýšené hladiny cirkulujících TFH lymfocytů. U těchto autoimunitních onemocněních může aberantní vývoj TFH lymfocytů na rozdíl od deficitů v protilátkové odpovědi naopak vést k produkci vysoce afinitních autoprotilátek proti vlastním tkáním, které pak mohou přispívat k rozsáhlým orgánovým poškozením či tvorbě a hromadění imunokomplexů v těle pacientů.[14]
Alternativní CD8+ TFC lymfocyty u chronických virových infekcí[editovat | editovat zdroj]
B buněčné zóny lymfoidních folikulů mohou sloužit jako rezervoár chronických virových infekcí, jelikož zde nejsou přítomné všechny složky antivirové obrany, a právě buňky které se zde nachází mohou být cílem virů (např. TFH lymfocyty pro virus HIV, B lymfocyty pro virus EBV). Bylo zjištěno, že v takovýchto situacích chronických virových infekcí mohou hrát důležitou roli tzv. folikulární cytotoxické lymfocyty TFC exprimující koreceptor CD8 společně se znaky TFH linie jako je exprese CXCR5 a Bcl6[15]. Nicméně pro přesný popis funkce těchto TFC buněk bude nutné další studium.[16]
Reference[editovat | editovat zdroj]
- ↑ BREITFELD, Dagmar; OHL, Lars; KREMMER, Elisabeth. Follicular B Helper T Cells Express Cxc Chemokine Receptor 5, Localize to B Cell Follicles, and Support Immunoglobulin Production. Journal of Experimental Medicine. 2000-12-04, roč. 192, čís. 11, s. 1545–1552. Dostupné online [cit. 2022-05-25]. ISSN 0022-1007. DOI 10.1084/jem.192.11.1545. PMID 11104797. (anglicky)
- ↑ a b CROTTY, Shane. Follicular Helper CD4 T Cells (T FH ). Annual Review of Immunology. 2011-04-23, roč. 29, čís. 1, s. 621–663. Dostupné online [cit. 2022-05-25]. ISSN 0732-0582. DOI 10.1146/annurev-immunol-031210-101400. (anglicky) Archivováno 25. 5. 2022 na Wayback Machine.
- ↑ HOŘEJŠÍ, Václav; BRDIČKA, Tomáš. Kdo pomáhá B-lymfocytům produkovat protilátky?. Vesmír. 2013-10-03, roč. 2013/10, čís. Vesmír 92, s. 542–544. Dostupné online.
- ↑ MA, Cindy S.; DEENICK, Elissa K.; BATTEN, Marcel. The origins, function, and regulation of T follicular helper cells. Journal of Experimental Medicine. 2012-07-02, roč. 209, čís. 7, s. 1241–1253. Dostupné online [cit. 2022-05-25]. ISSN 0022-1007. DOI 10.1084/jem.20120994. PMID 22753927.
- ↑ a b KUROSAKI, Tomohiro; KOMETANI, Kohei; ISE, Wataru. Memory B cells. Nature Reviews Immunology. 2015-03, roč. 15, čís. 3, s. 149–159. Dostupné online [cit. 2022-05-25]. ISSN 1474-1733. DOI 10.1038/nri3802. (anglicky)
- ↑ NUTT, Stephen L; TARLINTON, David M. Germinal center B and follicular helper T cells: siblings, cousins or just good friends?. Nature Immunology. 2011-06, roč. 12, čís. 6, s. 472–477. Dostupné online [cit. 2022-05-25]. ISSN 1529-2908. DOI 10.1038/ni.2019. (anglicky)
- ↑ MINTZ, Michelle A.; CYSTER, Jason G. T follicular helper cells in germinal center B cell selection and lymphomagenesis. Immunological Reviews. 2020-07, roč. 296, čís. 1, s. 48–61. Dostupné online [cit. 2022-05-25]. ISSN 0105-2896. DOI 10.1111/imr.12860. PMID 32412663. (anglicky)
- ↑ a b QI, Hai. T follicular helper cells in space-time. Nature Reviews Immunology. 2016-10, roč. 16, čís. 10, s. 612–625. Dostupné online [cit. 2022-05-25]. ISSN 1474-1733. DOI 10.1038/nri.2016.94. (anglicky)
- ↑ a b CROTTY, Shane. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity. 2014-10-16, roč. 41, čís. 4, s. 529–542. PMID: 25367570. Dostupné online [cit. 2022-05-25]. ISSN 1074-7613. DOI 10.1016/j.immuni.2014.10.004. PMID 25367570. (English)
- ↑ JI, Long-Shan; SUN, Xue-Hua; ZHANG, Xin. Mechanism of Follicular Helper T Cell Differentiation Regulated by Transcription Factors. Journal of Immunology Research. 2020-07-20, roč. 2020, s. 1–9. Dostupné online [cit. 2022-05-25]. ISSN 2314-8861. DOI 10.1155/2020/1826587. PMID 32766317. (anglicky)
- ↑ MA, Xiaoxue; NAKAYAMADA, Shingo; WANG, Jun. Multi-Source Pathways of T Follicular Helper Cell Differentiation. Frontiers in Immunology. 2021-02-25, roč. 12, s. 621105. Dostupné online [cit. 2022-05-25]. ISSN 1664-3224. DOI 10.3389/fimmu.2021.621105. PMID 33717120.
- ↑ a b TANGYE, Stuart G.; MA, Cindy S.; BRINK, Robert. The good, the bad and the ugly — TFH cells in human health and disease. Nature Reviews Immunology. 2013-06, roč. 13, čís. 6, s. 412–426. Dostupné online [cit. 2022-05-25]. ISSN 1474-1733. DOI 10.1038/nri3447. (anglicky)
- ↑ PALLIKKUTH, Suresh; PARMIGIANI, Anita; SILVA, Sandra Y. Impaired peripheral blood T-follicular helper cell function in HIV-infected nonresponders to the 2009 H1N1/09 vaccine. Blood. 2012-08-02, roč. 120, čís. 5, s. 985–993. Dostupné online [cit. 2022-05-25]. ISSN 0006-4971. DOI 10.1182/blood-2011-12-396648. PMID 22692510. (anglicky)
- ↑ CRAFT, Joseph E. Follicular helper T cells in immunity and systemic autoimmunity. Nature Reviews Rheumatology. 2012-06, roč. 8, čís. 6, s. 337–347. Dostupné online [cit. 2022-05-25]. ISSN 1759-4790. DOI 10.1038/nrrheum.2012.58. PMID 22549246. (anglicky)
- ↑ LEONG, Yew Ann; CHEN, Yaping; ONG, Hong Sheng. CXCR5+ follicular cytotoxic T cells control viral infection in B cell follicles. Nature Immunology. 2016-10, roč. 17, čís. 10, s. 1187–1196. Dostupné online [cit. 2022-05-25]. ISSN 1529-2908. DOI 10.1038/ni.3543. (anglicky)
- ↑ MA, Cindy S.; TANGYE, Stuart G. Cytotoxic T cells that escape exhaustion. Nature. 2016-09-15, roč. 537, čís. 7620, s. 312–314. Dostupné online [cit. 2022-05-25]. ISSN 0028-0836. DOI 10.1038/nature19428. (anglicky)